Recombinant Mouse Histone acetyltransferase p300 (Ep300), partial
-
中文名稱:小鼠Ep300重組蛋白
-
貨號(hào):CSB-YP007702MO
-
規(guī)格:
-
來源:Yeast
-
其他:
-
中文名稱:小鼠Ep300重組蛋白
-
貨號(hào):CSB-EP007702MO-B
-
規(guī)格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:小鼠Ep300重組蛋白
-
貨號(hào):CSB-BP007702MO
-
規(guī)格:
-
來源:Baculovirus
-
其他:
-
中文名稱:小鼠Ep300重組蛋白
-
貨號(hào):CSB-MP007702MO
-
規(guī)格:
-
來源:Mammalian cell
-
其他:
產(chǎn)品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:Ep300
-
Uniprot No.:
-
別名:Ep300; P300Histone acetyltransferase p300; p300 HAT; EC 2.3.1.48; E1A-associated protein p300; Histone butyryltransferase p300; EC 2.3.1.-; Histone crotonyltransferase p300; EC 2.3.1.-; Protein 2-hydroxyisobutyryltransferase p300; EC 2.3.1.-; Protein propionyltransferase p300; EC 2.3.1.-
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Partial
-
蛋白標(biāo)簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產(chǎn)品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復(fù)溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲(chǔ)存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質(zhì)期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項(xiàng):Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產(chǎn)品評(píng)價(jià)

相關(guān)產(chǎn)品
靶點(diǎn)詳情
-
功能:Functions as histone acetyltransferase and regulates transcription via chromatin remodeling. Acetylates all four core histones in nucleosomes. Histone acetylation gives an epigenetic tag for transcriptional activation. Mediates cAMP-gene regulation by binding specifically to phosphorylated CREB protein. Mediates acetylation of histone H3 at 'Lys-122' (H3K122ac), a modification that localizes at the surface of the histone octamer and stimulates transcription, possibly by promoting nucleosome instability. Mediates acetylation of histone H3 at 'Lys-27' (H3K27ac). Also functions as acetyltransferase for non-histone targets, such as ALX1, HDAC1, PRMT1 or SIRT2. Acetylates 'Lys-131' of ALX1 and acts as its coactivator. Acetylates SIRT2 and is proposed to indirectly increase the transcriptional activity of TP53 through acetylation and subsequent attenuation of SIRT2 deacetylase function. Acetylates HDAC1 leading to its inactivation and modulation of transcription. Acetylates 'Lys-247' of EGR2. Acts as a TFAP2A-mediated transcriptional coactivator in presence of CITED2. Plays a role as a coactivator of NEUROD1-dependent transcription of the secretin and p21 genes and controls terminal differentiation of cells in the intestinal epithelium. Promotes cardiac myocyte enlargement. Can also mediate transcriptional repression. Acetylates FOXO1 and enhances its transcriptional activity. Acetylates BCL6 wich disrupts its ability to recruit histone deacetylases and hinders its transcriptional repressor activity. Participates in CLOCK or NPAS2-regulated rhythmic gene transcription; exhibits a circadian association with CLOCK or NPAS2, correlating with increase in PER1/2 mRNA and histone H3 acetylation on the PER1/2 promoter. Acetylates MTA1 at 'Lys-626' which is essential for its transcriptional coactivator activity. Acetylates XBP1 isoform 2; acetylation increases protein stability of XBP1 isoform 2 and enhances its transcriptional activity. Acetylates PCNA; acetylation promotes removal of chromatin-bound PCNA and its degradation during nucleotide excision repair (NER). Acetylates MEF2D. Acetylates and stabilizes ZBTB7B protein by antagonizing ubiquitin conjugation and degradation, this mechanism may be involved in CD4/CD8 lineage differentiation. Acetylates GABPB1, impairing GABPB1 heterotetramerization and activity. In addition to protein acetyltransferase, can use different acyl-CoA substrates, such as (2E)-butenoyl-CoA (crotonyl-CoA), butanoyl-CoA (butyryl-CoA), 2-hydroxyisobutanoyl-CoA (2-hydroxyisobutyryl-CoA), lactoyl-CoA or propanoyl-CoA (propionyl-CoA), and is able to mediate protein crotonylation, butyrylation, 2-hydroxyisobutyrylation, lactylation or propionylation, respectively. Acts as a histone crotonyltransferase; crotonylation marks active promoters and enhancers and confers resistance to transcriptional repressors. Histone crotonyltransferase activity is dependent on the concentration of (2E)-butenoyl-CoA (crotonyl-CoA) substrate and such activity is weak when (2E)-butenoyl-CoA (crotonyl-CoA) concentration is low. Also acts as a histone butyryltransferase; butyrylation marks active promoters. Catalyzes histone lactylation in macrophages by using lactoyl-CoA directly derived from endogenous or exogenous lactate, leading to stimulates gene transcription. Acts as a protein-lysine 2-hydroxyisobutyryltransferase; regulates glycolysis by mediating 2-hydroxyisobutyrylation of glycolytic enzymes. Functions as a transcriptional coactivator for SMAD4 in the TGF-beta signaling pathway. Acetylates PCK1 and promotes PCK1 anaplerotic activity. Acetylates RXRA and RXRG.
-
基因功能參考文獻(xiàn):
- Study in mouse model found that liver stiffness activates hepatic stellate cells differentiation into myofibroblasts, which required nuclear accumulation of p300. PMID: 29454793
- Data (including data from studies in knockout and transgenic mice) suggest that Ep300 and Crebbp are limiting cofactors for pancreatic islet development (including gene expression regulation and cell proliferation), and hence for postnatal glucose homeostasis, with some functional redundancy. (Ep300 = E1A binding protein p300; Crebbp = CREB binding protein) PMID: 29217654
- Here the authors report a lipopolysaccharide-induced NFkappaB enhanceosome in which TonEBP is required for the recruitment of p300. Increased expression of TonEBP enhances the NFkappaB activity and reduced TonEBP expression lowers it. PMID: 27118681
- Data show that LPS induces endoplasmic reticulum (ER) stress and P300 activity via the XBP1/IRE1 pathway. PMID: 28743992
- Loss of p300 expression is associated with leukemogenesis. PMID: 27881875
- UTX-MLL4-p300 transcriptional regulatory network establishing an "active enhancer landscape" and defines a detailed mechanism for the joint deposition of H3K4me1 and H3K27ac. PMID: 28732206
- Acetylation-dependent control of global poly(A) RNA degradation by CBP/p300 and HDAC1-HDAC2 has been described. PMID: 27635759
- Data, including data from studies in cells from knockout mice, suggest that Prmt1 activity was necessary for c-Myc binding to acetyltransferase p300 in myeloid cells; Prmt1 inhibition decreases p300 recruitment to c-Myc target promoters and increased Hdac1 recruitment. [Prmt1, protein arginine N-methyltransferase 1; c-Myc = Proto-Oncogene Proteins c-myc; Hdac1 = histone deacetylase 1] PMID: 28652407
- In line with the acetyltransferase activity of p300, H3K27 acetylation was reduced after HDACi and resulted in the formation of heterochromatin in the PTGES1 gene. In conclusion, HDAC activity maintains PTGES1 expression by recruiting p300 to its gene PMID: 27913583
- 2-O, 3-O desulfated heparin inhibited HMGB1 release, at least in part, by direct molecular inhibition of p300 HAT activity. PMID: 27585400
- work shows that helenalin acetate inhibits C/EBPbeta by binding to the N-terminal part of C/EBPbeta, thereby disrupting the cooperation of C/EBPbeta with the co-activator p300. PMID: 27803164
- Findings indicate that E1a-binding protein (p300) is not required for the normal development or functioning of skeletal muscle. PMID: 26712218
- Hdac3 cooperates with p300 to prime and maintain oligodendrocyte identity PMID: 26859354
- p300 histone acetyltransferase activity is critical for Wnt-dependent palate mesenchymal cell proliferation and migration, both processes that play a significant role in morphogenesis of the palate. PMID: 26921506
- Data indicate that glucose-induced endothelial to mesenchymal transition in vivo and in vitro in the hearts of diabetic mice is possibly mediated by miR-200b and p300. PMID: 26718496
- This study demonstrated that p300 increase in skeletal muscle in muscle atrophy. PMID: 26372908
- This study demonstrates that p300 mediates histone acetylation of PS1 and BACE1 in a cellular model of Alzheimer's disease. PMID: 25051175
- p300 regulates a number of critical signaling pathways that control liver functions. PMID: 26100016
- p300 conditional knockout in Treg cells leads to minimal changes in Treg functions. PMID: 24835996
- on Bcl6, p300 functioned as a repressor and inhibited in conjunction with STAT5 or BCL6. PMID: 25088465
- These results suggest that the aristolochic acid dose dependently contributed to the development of nephropathy, and HDAC1 and p300 participate in the modulation of TGF-beta/Smad pathway-mediated renal interstitial fibrosis. PMID: 24796935
- ATF3 interacts with PPARgamma and represses PPARgamma-mediated transactivation through suppression of p300-stimulated coactivation in 3T3-L1 cells PMID: 25446101
- The formation of p300/CBP-mediated looping chromatin structures may recruit distal enhancers to create a concentration of factors for the transcription activation of genes that are involved in self-renewal and pluripotency. PMID: 24648406
- we demonstrated that inhibition of the acetylase function of p300 reduces both cell count and invasion in LM3 murine breast cancer cell line , and decreases tumor progression in the animal model PMID: 25240203
- This study revealed the occurrence of a tightly regulated Cbp/p300-dependent gene expression programme that drives a specific metabolic state both in progenitor spermatogenic cells and in late transcriptionally active spermatids. PMID: 24522976
- glucose-induced EndMT in vitro and in vivo is possibly mediated through TGFbeta and regulated by miR-200b and p300. PMID: 25335984
- Results show that p300 forms a complex network along with CBP and other transcription factors and function within this complex as master regulators of Foxp3+ Treg generation and cell survival. PMID: 25154413
- these results suggest that the KIX domains of CBP, and especially p300, are principal mediators of c-Myb-dependent gene activation and repression that is required for definitive hematopoiesis. PMID: 24340053
- a mutant mouse model demonstrates how c-Myb with p300 is required for oncogene-induced acute myeloid leukemia PMID: 24596419
- These results suggest that p300 and CBP play essential roles in maintaining photoreceptor-specific structure, function and gene expression. PMID: 23922782
- depletion of hepatic p300 reduces glycogen synthesis, decreases hepatic glycogen storage, and leads to relative hypoglycemia. PMID: 23770612
- Two independent regions of simian virus 40 T antigen increase CEBP/p300 levels and alter patterns of cellular histone acetylation. PMID: 24089570
- p300 is important for Foxp3(+) T(reg) cell function and homeostasis in vivo and in vitro. PMID: 23955711
- The p300 regulates GI differentiation primarily through Myb, rather than CREB. PMID: 23618903
- the role of Mdm2's RING domain in the control of p53 PMID: 22666487
- Studies demonstrate for the first time that PDGF-BB-induced TRAIL transcriptional activity requires the cooperation of Sp1, ac-H3 and p300, mediating increased expression of TRAIL which is important for VSMC proliferation and migration. PMID: 22415975
- p300 and PCAF cooperate in the control of microRNA 200c/141 transcription and epithelial characteristics PMID: 22384255
- Curcumin enhances the efficacy of chemotherapy by tailoring p65NFkappaB-p300 cross-talk in favor of p53-p300 in breast cancer PMID: 22013068
- mice in which the CBP CH1 domain structure is disrupted by deleting residues 342-393 are lean and insulin sensitized, as are p300 mutants PMID: 21803292
- Data support the acetyltransferase activities of p300/CBP in regulating FOXO signaling in skeletal muscle and suggests that acetylation may be an important mechanism to differentially regulate the FOXO homologues PMID: 21389279
- transcriptional coactivator p300 may be partially degraded during SMC differentiation, leaving an activated subpopulation with increased HAT activity and SMC differentiation-gene specificity PMID: 21179216
- p300 acetylates Pax5 and strongly enhances Pax5-mediated transcriptional activity PMID: 21357426
- Spatial conformation of chromatin in the osteopontin promoter leads to recruitment of coactivator p300, formation of a DNA looping complex with NF-kappaB and AP-1, and induction of osteopontin expression. PMID: 21257959
- These findings indicate possible mechanisms by which CBP/p300 tissue-specifically acts cooperatively with pCAF and HDAC3 either as a co-activator or co-repressor, respectively, for CLOCK/BMAL1. PMID: 19922678
- impairment of Lck-mediated CD4 coreceptor signaling by Nef is an important in vivo mechanism of HIV-1 pathogenesis PMID: 20810990
- Both TAZ domains of CREB-binding proteins CBP and p300 in the presence of stoichiometric concentrations of zinc adopt a well-defined structure in solution in the absence of binding partners. PMID: 15641773
- contribution of p300 to Rubinstein-Taybi syndrome (RSTS); analysis demonstrates p300(+/-) mice exhibit phenotypes reminiscent of RSTS patients but indicate activity of p300 in cognition likely less relevant or more susceptible to compensation than CBP PMID: 19822209
- KIX or CH1 domain deletion caused profound hematopoiesis defects. Losing other domains had only lineage-restricted effects. Mutation of the histone acetyltransferase domain had few effects on hematopoiesis & increased progenitor/stem-cell proliferation. PMID: 19822904
- conserved domains in two highly related coactivators (p300 and CBP) have contrasting roles in haematopoiesis PMID: 12384703
- has a distinct role in hamatopoietic stem cell self-renewal PMID: 12397173
顯示更多
收起更多
-
亞細(xì)胞定位:Cytoplasm. Nucleus.
-
數(shù)據(jù)庫鏈接:
Most popular with customers
-
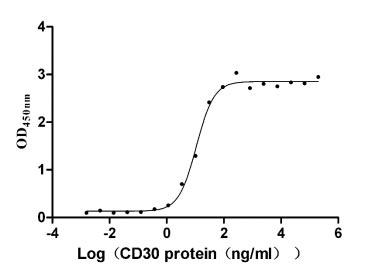
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Human Tumor necrosis factor ligand superfamily member 9 (TNFSF9), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-

Recombinant Macaca mulatta Semaphorin-4D isoform 1 (SEMA4D), partial (Active)
Express system: Mammalian cell
Species: Macaca mulatta (Rhesus macaque)
-
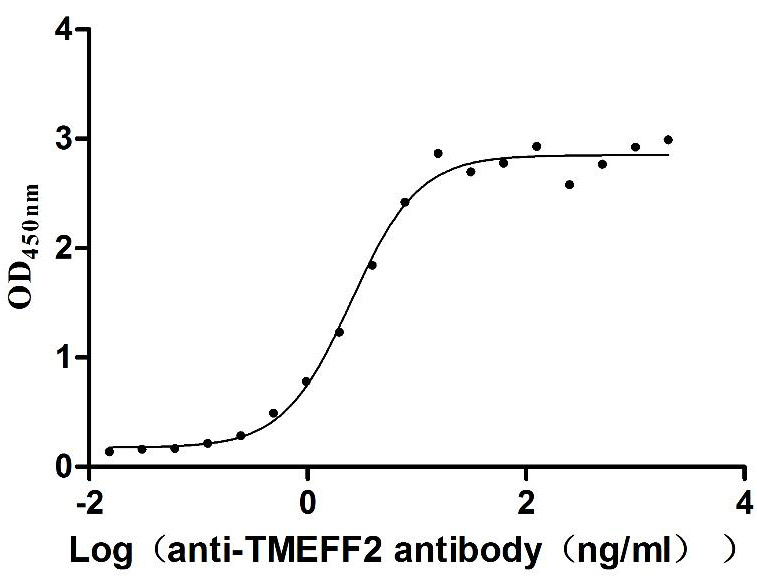
Recombinant Human Tomoregulin-2 (TMEFF2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
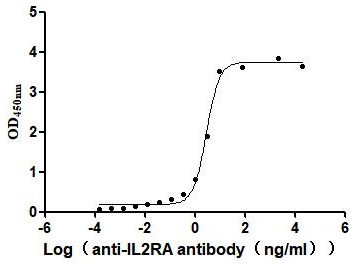
Recombinant Human Interleukin-2 receptor subunit alpha (IL2RA), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Mouse CUB domain-containing protein 1 (Cdcp1), partial (Active)
Express system: Mammalian cell
Species: Mus musculus (Mouse)
-
Recombinant Human Myosin regulatory light chain 12B(MYL12B) (Active)
Express system: E.coli
Species: Homo sapiens (Human)
-
Recombinant Human Kidney-associated antigen 1(KAAG1) (Active)
Express system: Baculovirus
Species: Homo sapiens (Human)